新發(fā)和再發(fā)傳染病構(gòu)成的威脅突出表明需要制定新的策略來克服這些感染�。此外,耐多藥微生物的出現(xiàn)促使我們需要開發(fā)標(biāo)準(zhǔn)抗生素方案的替代策略。其中一種策略是使用可同時(shí)發(fā)揮預(yù)防和治療作用的人單克隆抗體(Mab)。在過去的十年中�����,隨著mAb在該領(lǐng)域的廣泛應(yīng)用�����,包括感染性疾病在內(nèi)的廣泛病理領(lǐng)域的mAb試驗(yàn)迅速增加��。噬菌體展示�����、酵母展示���、mRNA展示����、核糖體展示����、細(xì)菌展示、哺乳動(dòng)物細(xì)胞表面展示等抗體展示技術(shù)的發(fā)展��,促進(jìn)了新型單克隆抗體的快速發(fā)展�����。這些體外選擇技術(shù)的基本原理源于表型(顯示的抗體結(jié)構(gòu))和基因型(抗體基因)之間的物理聯(lián)系����。即使有不同的展示方法,噬菌體展示仍被廣泛認(rèn)為是抗體展示的首選方式��。
成功的單抗選擇過程的一般要求是存在多種可供選擇的抗體基因組合庫。這種抗體基因的集合通常被稱為抗體文庫���?��?贵w文庫可以通過展示的抗體基因的來源來區(qū)分。這包括初始(健康個(gè)體)�����、免疫(受感染或免疫的個(gè)體)���、合成(化學(xué)合成)和半合成(天然免疫基因和化學(xué)合成基因的混合物)文庫���。人類抗體文庫的開發(fā)需要從人類供體中獲得B細(xì)胞���,由于所需樣品的特定特征�,如B細(xì)胞來源�,以及在研究中使用人類生物樣品方面的嚴(yán)格規(guī)定,有時(shí)會(huì)成為瓶頸�。除人類外,動(dòng)物也是生成抗體文庫的B細(xì)胞的重要來源�����。在傳染病的背景下,免疫文庫是非常有吸引力的選擇���,因?yàn)樗鼈儽辉O(shè)計(jì)用來反映受感染個(gè)體或免疫動(dòng)物的免疫反應(yīng)���,反映針對(duì)該感染的有偏見的抗體基因庫。這只在B細(xì)胞能夠引起對(duì)感染的免疫反應(yīng)的情況下是正確的����。因此,對(duì)特定感染的免疫反應(yīng)的性質(zhì)是免疫抗體文庫設(shè)計(jì)的關(guān)鍵���,并決定了文庫的效用���、質(zhì)量和多功能性。然而��,通過免疫球蛋白基因組合混合構(gòu)建的體外免疫抗體庫庫可能不能完全反映天然抗體庫的真實(shí)性質(zhì)����,因?yàn)橹劓満洼p鏈的隨機(jī)配對(duì)可能導(dǎo)致折疊錯(cuò)誤的無功能抗體克隆。在這里,我們從免疫文庫的角度回顧了對(duì)不同感染的免疫反應(yīng)如何影響抗感染的鑒定�,并改進(jìn)了免疫抗體文庫用于噬菌體展示的庫表示的考慮。
免疫抗體庫
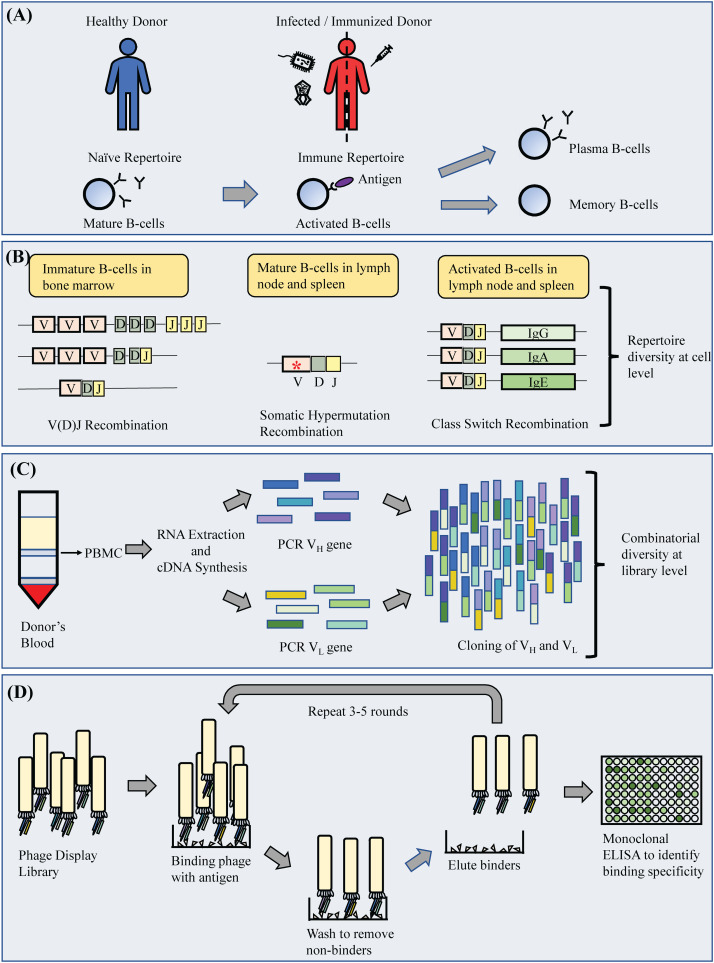
B細(xì)胞作為免疫系統(tǒng)的主要組成部分�����,保護(hù)我們的身體免受有害抗原的影響��,在感染發(fā)生之前和之后不斷產(chǎn)生抗體����。健康狀態(tài)下的抗體庫具有足夠的多樣性,可以產(chǎn)生針對(duì)新感染的應(yīng)答�,也可以記住舊感染。B細(xì)胞的這一主要特征是通過兩個(gè)主要機(jī)制實(shí)現(xiàn)的�,即變量(V)的V(D)J重組、多樣性(D)和連接(J)基因片段和體細(xì)胞超突變(SHM)����。這包括在較小程度上增加抗體組庫多樣性的非常規(guī)次要機(jī)制,包括違反12/23重組規(guī)則的非標(biāo)準(zhǔn)重組����、shm相關(guān)的基因插入和缺失��、抗體的非互補(bǔ)決定區(qū)(non- cdr)與抗原的直接接觸、翻譯后修飾��、構(gòu)象異質(zhì)性和使用非蛋白輔因子���。這些機(jī)制共同導(dǎo)致抗體cdr的不同變化�,形成主要抗原結(jié)合位點(diǎn)���。導(dǎo)致成熟抗體基因產(chǎn)生的事件也是多方面的�����,因?yàn)槎鄠€(gè)可變基因的重組一開始就提供了很大的組合多樣性�����,并進(jìn)一步擴(kuò)展為不同的重鏈(VH)和輕鏈(VL)組合(圖1)����。由于SHM和相關(guān)機(jī)制在遇到抗原時(shí)被引出�,暴露于感染會(huì)影響最終產(chǎn)生的抗體庫,因?yàn)榭贵w庫會(huì)發(fā)生偏倚�,并被塑造成與入侵病原體作戰(zhàn)的抗體庫。
Fig. 1. Schematic diagram of monoclonal antibody generation from sampling to isolation of antibodies.
盡管免疫抗體庫在分離疾病特異性結(jié)合物方面提供了更好的前景�,但目前的大多數(shù)噬菌體展示抗體方法主要集中在使用初始細(xì)胞庫來制備抗傳染病的單克隆抗體,正如美國(guó)食品藥品監(jiān)督管理局(FDA)和歐洲藥品管理局(EMA)批準(zhǔn)的大多數(shù)噬菌體展示單克隆抗體。
免疫抗體庫在傳染病中的應(yīng)用
源自免疫抗體庫的抗體具有用作診斷試劑或治療劑的潛力�。雖然從康復(fù)患者創(chuàng)建的免疫文庫是治療性mAb的理想選擇,但也可以從免疫動(dòng)物(如小鼠�、雞、美洲駝�����、羊駝�����、駱駝��、綿羊����、鯊魚和非人靈長(zhǎng)類動(dòng)物)中創(chuàng)建免疫文庫。
細(xì)菌感染
從歷史上看�����,細(xì)菌感染用動(dòng)物的抗血清治療��,后來用抗生素治療�。細(xì)菌的進(jìn)化導(dǎo)致了耐藥菌株的激增,這削弱了基于抗生素的治療策略的療效���。因此�,mAb被吹捧為抗生素耐藥細(xì)菌感染的潛在替代品�����。一般來說��,細(xì)菌毒素在增強(qiáng)感染方面起著重要作用�����。破傷風(fēng)類毒素免疫文庫是首批由免疫人外周B細(xì)胞庫構(gòu)建的組合免疫抗體庫之一�����,從中成功分離出多個(gè)mAb����,但未評(píng)估其中和潛力。
病毒感染
免疫抗體庫也被廣泛用于發(fā)現(xiàn)針對(duì)病毒感染的中和單克隆抗體�����,如登革熱、埃博拉病毒病�����、乙型肝炎�����、人類免疫缺陷病毒(HIV)感染��、流感���、麻疹���、狂犬病和呼吸道合胞病毒(RSV)。大多數(shù)商業(yè)化抗病毒單抗開發(fā)的共同和有效策略是在感染的初始階段通過與病毒表面受體(如包膜糖蛋白(gp)�����、刺突蛋白和受體結(jié)合域(RBD))的抗體結(jié)合使病毒失活�����,從而抑制病毒進(jìn)入宿主細(xì)胞的內(nèi)化����。免疫抗體庫中抗體庫的多樣性也可用于廣譜中和單克隆抗體的分離�。對(duì)來自免疫抗體庫的單克隆抗體進(jìn)行的研究表明�,單克隆抗體對(duì)來自同一家族或亞型的多種病毒的保守表位具有交叉反應(yīng)性�����,這一點(diǎn)在一項(xiàng)關(guān)于H5N1流感的研究中得到了證明����,從H5N1流感幸存者創(chuàng)建的組合免疫庫中分離出的300多種單克隆抗體能夠中和H1和H5亞型流感病毒。這說明了組合免疫文庫在選擇針對(duì)攜帶相似表位的密切相關(guān)抗原的抗體方面的應(yīng)用��。
寄生蟲感染
由原生動(dòng)物�����、蠕蟲和體表寄生蟲引起的寄生蟲病也是一個(gè)主要的衛(wèi)生保健負(fù)擔(dān)��。最常見的例子之一是引起瘧疾的蚊媒原生動(dòng)物瘧原蟲�����。已經(jīng)建立了免疫文庫�,用于分離針對(duì)惡性瘧原蟲Pfs48/45配子表面蛋白和惡性瘧原蟲裂殖子表面蛋白-1 Block 2區(qū)以及其他寄生蟲如豬帶絳蟲Ts14糖蛋白和剛地弓形蟲MIC2蛋白的單克隆抗體�����。只有針對(duì)惡性瘧原蟲配子細(xì)胞表面蛋白的單克隆抗體阻斷了吸血后瘧原蟲在蚊子體內(nèi)的發(fā)育�。這表明了免疫文庫衍生抗體在阻斷寄生蟲病傳播方面的潛在用途��。隨著單克隆抗體發(fā)現(xiàn)技術(shù)的成熟和更多單克隆抗體的出現(xiàn)����,有望為無有效藥物治療的感染性疾病提供更經(jīng)濟(jì)有效的解決方案。
免疫抗體文庫不僅能夠產(chǎn)生高親和力的疾病特異性單克隆抗體����,而且還具有產(chǎn)生針對(duì)其他靶蛋白的單克隆抗體的多樣性,使其成為抗體噬菌體展示實(shí)驗(yàn)室中初始細(xì)胞文庫不可缺少的替代品�。
參考資料
[
1]H.D. Marston, Monoclonal antibodies for emerging infectious diseases — borrowing from history
,N. Engl. J. Med., 378 (16) (2018), pp. 1469-1472
[2]E.K. Wagner,Engineering therapeutic antibodies to combat infectious diseases
,Curr. Opin. Chem. Eng., 19 (2018), pp. 131-141
[
3]K.B. Hoehn,The diversity and molecular evolution of B-cell receptors during infection
,Mol. Biol. Evol., 33 (5) (2016), pp. 1147-1157
[
4]A. Kanyavuz,Breaking the law: unconventional strategies for antibody diversification
,Nat. Rev. Immunol., 19 (6) (2019), pp. 355-368
[
5]L.M. Walker, D.R. Burton
,Passive immunotherapy of viral infections: ‘super-antibodies’ enter the fray
,Nat. Rev. Immunol., 18 (5) (2018), pp. 297-308